






One, No One and One Hundred Thousand
During sea urchin fertilization, the activating spermatozoon triggers a series of morphological and physiological events, such as egg’s plasma membrane depolarization, initiation of calcium signals that are concomitant with the changes in the actin cytoskeleton dynamics1 and the elevation of the fertilization envelope (Figure 1), as a result of the cortical granules exocytosis2. It is widely accepted that the sodium-dependent rapid changes of the membrane potential induced by the spermatozoon upon its fusion with the egg, serves as a (fast block) mechanism to prevent the entry of additional spermatozoa3, whereas the elevation of the fertilization envelope has been suggested to work as a barrier to mechanically block their penetration (slow block)4. At variance with the prevailing view that places emphasis on the depolarization of the egg plasma membrane to prevent polyspermy,3 we have provided the evidence that reduction of the amount of external sodium in the seawater, where fresh intact eggs are inseminated, fails to induce the polyspermic fertilization as one would expect according to the current view (Figure 2). Furthermore, we have shown that the observed abnormal cleavage of eggs fertilized in low sodium (Figure 3) is due to alterations in actin filament dynamics and is not the result of the formation of multiple sperm asters. Our results have also shown, at the variance with the prevailing idea that the fertilization envelope, already shown to be too slow to act as a mechanical block to polyspermy, is inefficient to ensure monospermy even when it is fully elevated5,6. We thus suggest that, at fertilization, the fast dynamic structural change of the cortical actin cytoskeleton of the egg - not the plasma membrane depolarization - prevents the entry of additional spermatozoa. Our results are in agreement with the hypothesis put forward by Ernest Everett Just a century ago2, according to which the integrity and the changes of the structure of the “ectoplasm” (the outer region of the egg cytoplasm) are crucial for the normal response of the egg during the fertilization reaction2. In line with this, our Laboratory has consistently demonstrated that starfish and sea urchin eggs are penetrated by multiple spermatozoa when the actin cytoskeleton of the egg surface has been injured by agents affecting its structural organization and dynamics. Our results confirm the experimental observations of Ernest Everett Just, who wrote: "…Normally monospermic eggs can be rendered polyspermic by experimental treatment as staling (long residence in sea-water), treatment with alkali, acids or poisons, subjection to low temperature, in short, by methods for inducing injury or weakness… and … polyspermy in them thus is a sign of weakness and hence pathological7…"
References
1. Santella L, Limatola N, Vasilev F, Chun J.T.(2018) Maturation and fertilization of echinoderm eggs: Role of actin cytoskeleton dynamics. Biochem. Biophys. Res. Commun. 506, 361-371.
2. Just E E. (1919) The fertilization-reaction in Echinarachnius Parma. I. Cortical response of the egg to insemination. Biol. Bull. 36, 1-10.
3. Jaffe L A. (1976). Fast block to polyspermy in sea urchin eggs is electrically mediated. Nature 261, 68-71.
4. Gilbert, Scott F. (2014) Developmental Biology, 10th ed. Sunderland, MA USA: Sinauer Associates, Inc. Publishers. p. 129.
5. Limatola N, Vasilev F, Chun J T, Santella L. (2019). Sodium-mediated fast electrical depolarization does not prevent polyspermic fertilization in Paracentrotus lividus eggs. Zygote 27, 241-249. page 1 of 9.
6. Dale B. (2019) New experimental data refuting the idea of a fast electrical block to polyspermy in sea urchin eggs. Zygote. 27, 193-194.
7. Just E E. (1939) The Biology of the Cell Surface. P. Blakiston’s Son & Co., Philadelphia, USA.
 Figure 1. Cortical reaction in an intact Paracentrotus lividus egg. Bright field view (left panel) and scanning electron micrographs (right panel) of the fertilized eggs at the key moments of the cortical reaction (a and b). The contact of the fertilizing spermatozoon with the egg plasma membrane generates a dimple on the egg surface at which the fertilization envelope (FE) begins to elevate (c and d). A few seconds later, the concavity becomes more pronounced, while the spermatozoon penetrates deeper into the egg with the formation of the fertilization cone (arrow in e). After a few minutes, the spermatozoon completes its incorporation and the fertilization envelope is fully elevated over the entire surface of the egg (e and f). White arrows indicate the site of sperm-egg interaction and penetration. Scale bars = 10 µm.
Figure 1. Cortical reaction in an intact Paracentrotus lividus egg. Bright field view (left panel) and scanning electron micrographs (right panel) of the fertilized eggs at the key moments of the cortical reaction (a and b). The contact of the fertilizing spermatozoon with the egg plasma membrane generates a dimple on the egg surface at which the fertilization envelope (FE) begins to elevate (c and d). A few seconds later, the concavity becomes more pronounced, while the spermatozoon penetrates deeper into the egg with the formation of the fertilization cone (arrow in e). After a few minutes, the spermatozoon completes its incorporation and the fertilization envelope is fully elevated over the entire surface of the egg (e and f). White arrows indicate the site of sperm-egg interaction and penetration. Scale bars = 10 µm.
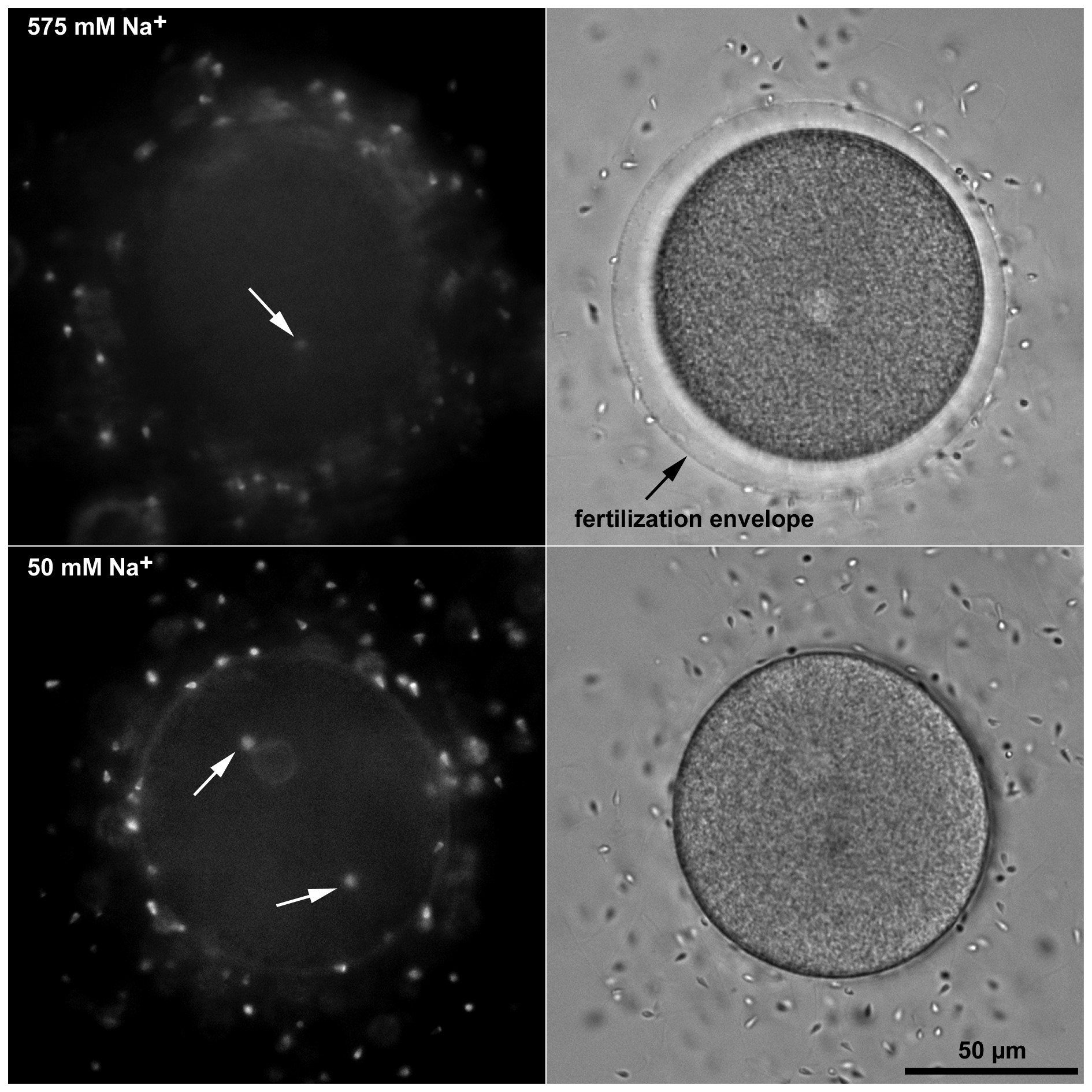 Figure 2. Fertilization in low sodium does not induce polyspermy. P. lividus eggs were transferred from natural to the artificial seawater containing various amount of Na+ and fertilized. In 50 mM Na+ the penetration of more than one sperm into the egg occurred very rarely. Interestingly, the spermatozoa stained with Hoechst 33342 always entered at opposite poles of the egg (arrows) arguing against the existence or effectiveness of a fast transmitting electrical mechanism that blocks polyspermy at the plasma membrane level. Elevation of the fertilization envelope is shown in the bright field view.
Figure 2. Fertilization in low sodium does not induce polyspermy. P. lividus eggs were transferred from natural to the artificial seawater containing various amount of Na+ and fertilized. In 50 mM Na+ the penetration of more than one sperm into the egg occurred very rarely. Interestingly, the spermatozoa stained with Hoechst 33342 always entered at opposite poles of the egg (arrows) arguing against the existence or effectiveness of a fast transmitting electrical mechanism that blocks polyspermy at the plasma membrane level. Elevation of the fertilization envelope is shown in the bright field view.
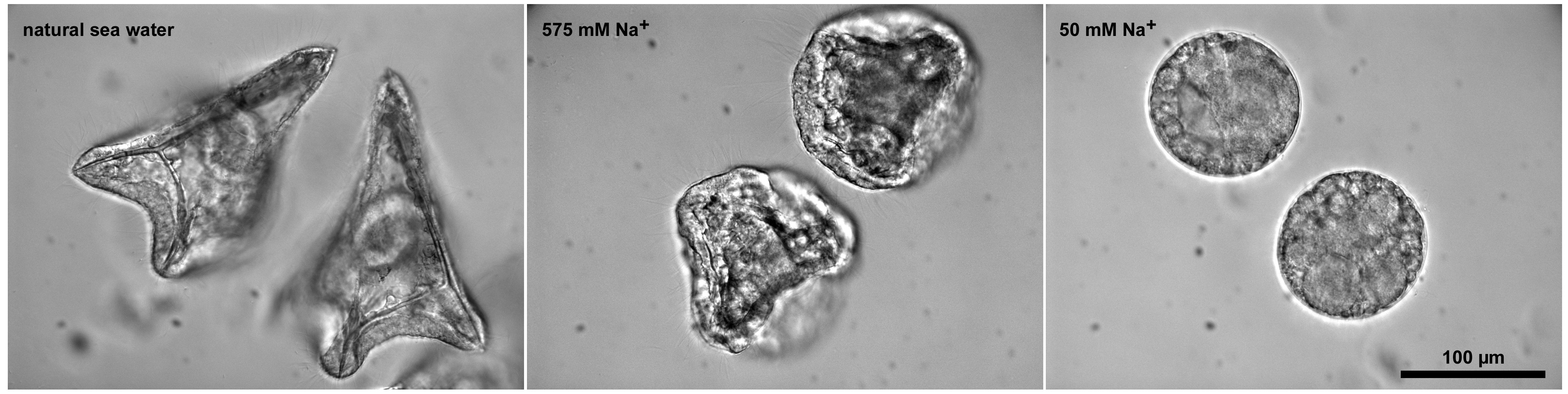 Figure 3. Embryonic development of monospermic sea urchin eggs fertilized in low Na+. Bright field view images of the embryos 1day after insemination.
Figure 3. Embryonic development of monospermic sea urchin eggs fertilized in low Na+. Bright field view images of the embryos 1day after insemination.